Biomass accumulation by Miscanthus in Illinois as evidence for the possibility of perennial grains?
This post was written in response to R. Ford Denison's review on the Darwinian Agriculture Blog of a paper in which my colleagues and I attempted to use a Maize vs. Miscanthus study ((Dohleman and Long, 2009) as evidence that herbaceous perennials theoretically could do everything that annual crops do (produce grain, leaves, roots) and have enough photosynthate left over to survive the winter as rhizomes.David Van Tassel, The Land Institute, 28 Nov 2012
***
Ford is not the first person to be underwhelmed by my Miscanthus vs. maize thought experiment. Maybe he (or other blog readers) can help me understand why it is not a useful case study for making the argument that herbaceous perennials could be bred to allocate more photosynthate to reproductive structures (e.g., seed) without sacrificing their ability to overwinter. The fact that the Miscanthus cultivar used in this study (Dohleman and Long, 2009) was a sterile triploid is a real barrier for many people and I've have a hard time understanding that.
First the things I do understand:
(a) "...it wasn't until the third year of growth that Miscanthus plants got big enough to capture that much sunlight and produce that much biomass." Good point. Yes. Perennials have a slower establishment phase. Many of the perennials we work with at The Land Institute do get big enough to flower in their first season, but their seedling relative growth rate is lower than their annual counterparts and peak yields are not realized until the second year. This is indeed a tradeoff and it is one that we have acknowledged. We point out that an establishment period of months to years has not prevented alfalfa, grass hay, grapes, blueberries, apples, oil palm, etc. from being viable and profitable crops. The establishment costs must be amortized over the productive lifespan of the stand and the final determination of profitability will depend on the yield and value of the crop and the expected stand lifespan. Canny farmers may find ways to reduce the establishment costs by grazing the young perennial or raising an annual nurse crop or intercrop. We don't know any of those things yet, and when we do we may find that perennial grains are not profitable, but at the moment we are considering whether they are possible.
(b) "...wouldn't growing Miscanthus year after year have similar risks?" [Similar to the risk of disease/pests in continuous corn]. Again, a good point. I agree that yearly crop rotation is not possible with perennial crops. However, again, we point out that there are already many perennial crops--just not perennial grains--and this challenge has not prevented their success. Much of the R&D investment in bioenergy crops has been on perennials such as Miscanthus, switchgrass, poplar, willow, bamboo, etc. Are these crops doomed? What about alfalfa and other hay crops? Forest plantations of yellow pine and Douglas fir? Doomed? If not, why not and what is the difference between a perennial fruit/hay/energy/timber crop and a perennial grain crop? Perhaps they are all doomed in the long term and we should expect that stands of all perennial crops will need to be replaced with different species or new disease resistant cultivars every 5-10 years. Furthermore, while Ford favors diversity in time (rotations) in his book, he acknowledges the potential to reduce pest and disease incidence by deploying diversity in space. Intra and interspecific diversity may prove important in moderating disease and herbivory in perennial grains.
Now onto the part of the thought experiment where I'm mystified by other people’s mystification. Ford’s response is typical: “What about seed yield? Zero. The genotype used doesn't make any seeds. It's propagated from cuttings. So while corn is recycling nitrogen from leaves to seeds in the autumn, Miscanthus just keeps growing. Ok, but weren't we talking about the potential of herbaceous perennials to have high grain yields?”
Background: we argued that the Dohleman and Long study was an excellent opportunity to compare the productivity of a perennial vs. and annual. Miscanthus and Zea are closely related genera with C4 photosynthesis. Both have been selected for high yield. Unlike many studies of species with different life histories, this experiment was done on a field scale with replicated plots and thorough characterization of the crops' photosynthesis throughout the season. We know of no other such experiment. The Miscanthus yielded about 10 Mg/ha more above-ground biomass than the maize each year. The extra biomass was attributed to earlier canopy development and later senescence--a longer growing season. This is exactly what we have been predicting for perennial grains since dormant rhizomes can break dormancy and rapidly produce leaves in cold soil much more quickly than can small dormant seeds.
Rationale: Why the interest in a thought experiment involving biomass accumulation rather than genetics and breeding? Because a common reaction to the proposal that perennials could be domesticated as grains is something along the lines of: "Well, perennial herbs are great at producing foliage and roots, but they don't produce much (if any) seed or fruit. If you force them to produce large quantities of reproductive biomass, they will have to rob either their root systems or leaves. This would turn them into annuals or perennials too weak and slow growing to producing high grain yields." Our position, argued in detail elsewhere (Van Tassel et al., 2010 and DeHaan et al., 2005) is that wild perennials invest heavily in other costly structures and functions that could be robbed to pay for increased seed production. We felt that this Miscanthus study supports that claim.
 Relevance: Dohleman and Long showed that both species intercepted a similar amount of PAR once their canopies were fully established, so it's not like the Miscanthus has a much larger or more efficient canopy than maize. Where is the extra 10 Mg/ha of above ground biomass going if there is no seed to fill? In a previous study (Beale and Long, 1997) showed that Miscanthus has a great deal of stem material--about 20 Mg/ha (2kg/m2), the vertical line-shaded area under the curve in their figure 1 (right), or approximately the same amount of stalk as maize stalks, leaves and ears combined. Thus, it seems fair to say that Miscanthus has much more stalk material than needed simply to hold up the canopy, since Maize can hold up its similar canopy with much less stalk biomass. Like woody plants, Miscanthus may invest in more stalk than needed for optimum light interception as a result of a history of competition with other tall grasses or shrubs. Similarly, each maize plant clearly invests more in seed biomass than needed to replace itself the following year; maize plants have a history of competition on the basis of seed p
Relevance: Dohleman and Long showed that both species intercepted a similar amount of PAR once their canopies were fully established, so it's not like the Miscanthus has a much larger or more efficient canopy than maize. Where is the extra 10 Mg/ha of above ground biomass going if there is no seed to fill? In a previous study (Beale and Long, 1997) showed that Miscanthus has a great deal of stem material--about 20 Mg/ha (2kg/m2), the vertical line-shaded area under the curve in their figure 1 (right), or approximately the same amount of stalk as maize stalks, leaves and ears combined. Thus, it seems fair to say that Miscanthus has much more stalk material than needed simply to hold up the canopy, since Maize can hold up its similar canopy with much less stalk biomass. Like woody plants, Miscanthus may invest in more stalk than needed for optimum light interception as a result of a history of competition with other tall grasses or shrubs. Similarly, each maize plant clearly invests more in seed biomass than needed to replace itself the following year; maize plants have a history of competition on the basis of seed pHere's how my mind works: as a thought experiment, it seems fair to imagine using breeding to develop plants with a different allocation ratio than their existing ratio. Changes in allocation, sometimes dramatic, happened during the domestication of all of our annual grain crops and the breeding done during the green revolution. As we simply explore whether perennial grains are possible, it seems fair to imagine substituting one kind of competitive structure with another. It seems fair to imagine replacing a portion of the stem biomass with sexually reproductive biomass. It seems fair because neither excess stem material nor ears are principally a photosynthetic structure and neither is required for supporting the function of the root or canopy systems.
Hypothesis: In order for perennial grains with yields similar to maize to be feasible, a perennial grass similar to maize should be able to produce (a) as much stem and leaf biomass as maize; (b) belowground storage sufficient for paying overwintering and re-growing costs ; (c) additional aboveground or belowground biomass equal to the amount of maize seed (biomass that could be reallocated to seeds). In short, we would want to see that a herbaceous perennial could each year produce at least an equal amount of removable aboveground biomass (including grain) as an annual without sacrificing the ability to survive and regrow.
My evaluation: The perennial grain feasibility hypothesis cannot be rejected on the basis of Miscanthus perfomance for the three tests listed above. Dohleman and Long did not measure belowground storage but plainly there was sufficient for survival and regrowth. In addition--as a bonus-- Miscanthus produces an extra 10 Mg of aboveground biomass that could be reallocated for anything. On top of that, there is almost certainly more rhizome production than really needed for overwintering (see the stippled area in the figure above showing a standing rhizome mass of 5-10 Mg/ha). To me, this shows that in theory perennial grains have the potential to have equal (or higher) yields than annual grains like maize. In reality, getting perennial plants to reallocate massively to sexual structures is a huge challenge. This is a genetic challenge not a physiological one. It may prove impossible. There may be serious ecological challenges as suggested above. Plainly there will be trade-offs: the domesticated perennials will be shorter and have less massive stalks. They will not reproduce asexually as aggressively. They will likely become very uncompetitive in their original wild habitat. These are acceptable tradeoffs.
However, as I suggested above, this argument has often proven to be unpersuasive and I'd appreciate feedback. Possible objections:
1. Genetic. People can't believe that there is sufficient genetic variation to allow selection for increased allocation to seed and decreased allocation to stem. To my mind, this is not relevant for a thought experiment. However, in case it matters, we should note that there are seed producing Miscanthus cultivars. I have seen them growing side by side with the sterile triploids. They look very similar and I would predict that they have similar biomass production. Unfortunately the yield data is proprietary as these are being bred by private companies as bioenergy crops. We have no reason to believe that there is no genetic variation for seed or stem traits in these sexual cultivars...not that anyone is talking about developing grain Miscanthus.
2. Construction costs. Perhaps people instinctively believe that investing in seeds is far more costly than stems, per unit of final biomass. If this is true I need to hear about it. My only source of information suggests that, if anything, stems are slightly more costly (Penning deVries et al., 1989).
3. Opportunity costs. We can all imagine that a plant going to seed should undergo programmed death in order to be able to remobilize carbon and nitrogen for seed filling (--but not for stalk construction?). I think that this is the argument Ford is making in his blog entry, but I'd like to hear whether this is a quantitative objection or a qualitative one:
(a) Quantitative: Are people thinking that programmed tiller death simply reduces the season-extension to which Dohleman and Long ascribed the greater productivity of Miscanthus? If this is the thought then I'm curious how much season extension it is thought would be enough to make perennial grains feasible? In 2008 Miscanthus accumulated the majority of its aboveground biomass by the same time that maize did, so there would seem to have been no penalty had tillers senesced during grain-filling in the perennial (see "Figure 1" below). In 2007, Miscanthus continued to accumulate biomass after maize maximum accumulation so there would have been a penalty. However, Miscanthus had already accumulated about 4 Mg/ha more than maize by September 7. Would an extra 4 Mg/ha of photosynthate be enough to permit overwintering?
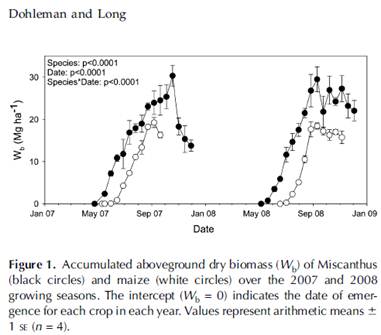
The extra aboveground biomass is a bonus and not required for the feasibility of perennial grains, however, perhaps people are thinking that the late season growth is required to refill the belowground stores. Programmed death of the shoots would deprive the rhizomes of this recharging period. So now the question becomes whether, assuming zero rhizome recharge during the spring (a very conservative assumption, since the figure above seems to show recharge of the rhizomes during the summer), the extra 4 Mg/ha of aboveground biomass, if reallocated belowground, would be enough for winter metabolism and spring regrowth. Unfortunately, no one knows how much of the Miscanthus rhizome biomass is needed for survival/regrowth and how much is for asexual reproduction. Can we make guesses about the minimum amount of biomass reallocation required?
- One estimate of the biomass required to support enough dormant meristems to form a full crop canopy is the amount of seed required to replant an annual crop. Seeds are a form of dormant meristem. They respire during the winter and during germination. Dohleman and Long planted 80,000 seeds/ha. Maize seeds are about 0.0004 kg, thus they planted about 32 kg/ha (0.032 Mg/ha).
- Presumably, seeds are a more efficient way to store meristems over the winter than rhizomes, but undersized for rapid canopy deployment. The question is how much more efficient and how undersized. Do rhizomes require twice this amount? Twenty times? Miscanthus in Illinois appears to have more than 100 times more “excess” early season productivity (4 vs. 0.032 Mg/ha) that could be used for this recharge.
- Theoretically, the roots and rhizomes themselves need not be reconstructed each year (for a perennial); only the nonstructural carbohydrates depleted in winter metabolism and spring growth need to be recharged. I haven’t been able to find useful values in the literature. Any ideas? Papers?
4. Asexual plants are fundamentally different. I suspect that this is closer to the true reason that some people aren't persuaded by the Miscanthus story. Again I can imagine different reasons for this but I none of them really make sense to me: (a) Asexual plants have experienced very different selection pressures than sexual plants. Selection for vegetative vigor necessarily reduces sexual fertility (negative genetic correlation), not just seed size or reproductive effort. (b) Recent triploids are something like cancer tumors. They may not have experienced generations of directional selection for vegetative vigor, but they have been released from developmental controls and are examples of unrestrained growth. They seem healthy to us, but from an evolutionary timescale, they are heading for extinction.
Is there a sense that a sexual species could never have experienced such strong selection--or release from developmental controls--permitting extreme photosynthetic or vegetative vigor and thus it is unrealistic to use the productivity of an asexual clone as any kind of guide to the possible phenotypes of sexual lineages? If this is how people feel about Miscanthus, it would be helpful to me to hear what the proposed mechanisms are. Why would high vegetative vigor be incompatible with sexuality?
5. I suspect there are other reasons people can offer to explain why the biomass accumulation of a sterile triploid cultivar should not be used to establish a ballpark annual biomass budget of a hypothetical herbaceous, C4 perennial grain, and to compare that budget with a related annual species. Or there may be other reasons why it is unreasonable to suggest that evolution or plant breeding can result in substantial changes in partitioning of the biomass between some sub-categories without affecting the bottom line. [It is certainly reasonable to assume that these changes will be difficult, costly, and dependent upon the genetics of particular candidate species.] Please comment. I want to know where my blind spots are.
